
外观
Lesson 14 Dynamics of biological networks (2)
生物网络:
- 直接物理交互:比如核糖体附着在 DNA 上
- 相关性:有相关性的不同分子或者细胞器
- ……
上面的这些内容整体上可以组成一个「图」,图中可以存在加权重的连接等等.
一维正反馈效应:
这个方程很明显难以得到解析的解,而且其解析解的实际意义也并不大. 事实上,我们更倾向于考虑分析这个方程的稳定性. 也就是对于 这样的情况,分析 造成的结果. 稳定态和不稳定态之间存在「分叉」.
提示
我认为,这里的一维方程分叉和理论力学混沌理论中的 Logistic 映射可能有类似的机制.
基因开关:考虑某个基因被抑制的概率为
(仍然取一个 Hill 反应的形式.) 转录对应的方程为
一共有两个基因,这是无量纲化的方程. 我们期望先找到不动点,也就是系统长时演化之后的情况;之后在不动点附近展开.
这个系数矩阵 (Jacobi Matrix) 给出两个本征值,它们的正负表现了两个不动点的稳定性,可能出现鞍点、旋进或者旋出的不同情况.
对于一个非常复杂的大型生物网络,我们的处理哲学是「Less is More」,期望达到的效果是把不重要的成分全部丢掉. 操作中类似于统计方法中的 - NN,用周围的状态给这个网格节点投票,决定其下一个步骤的演化.
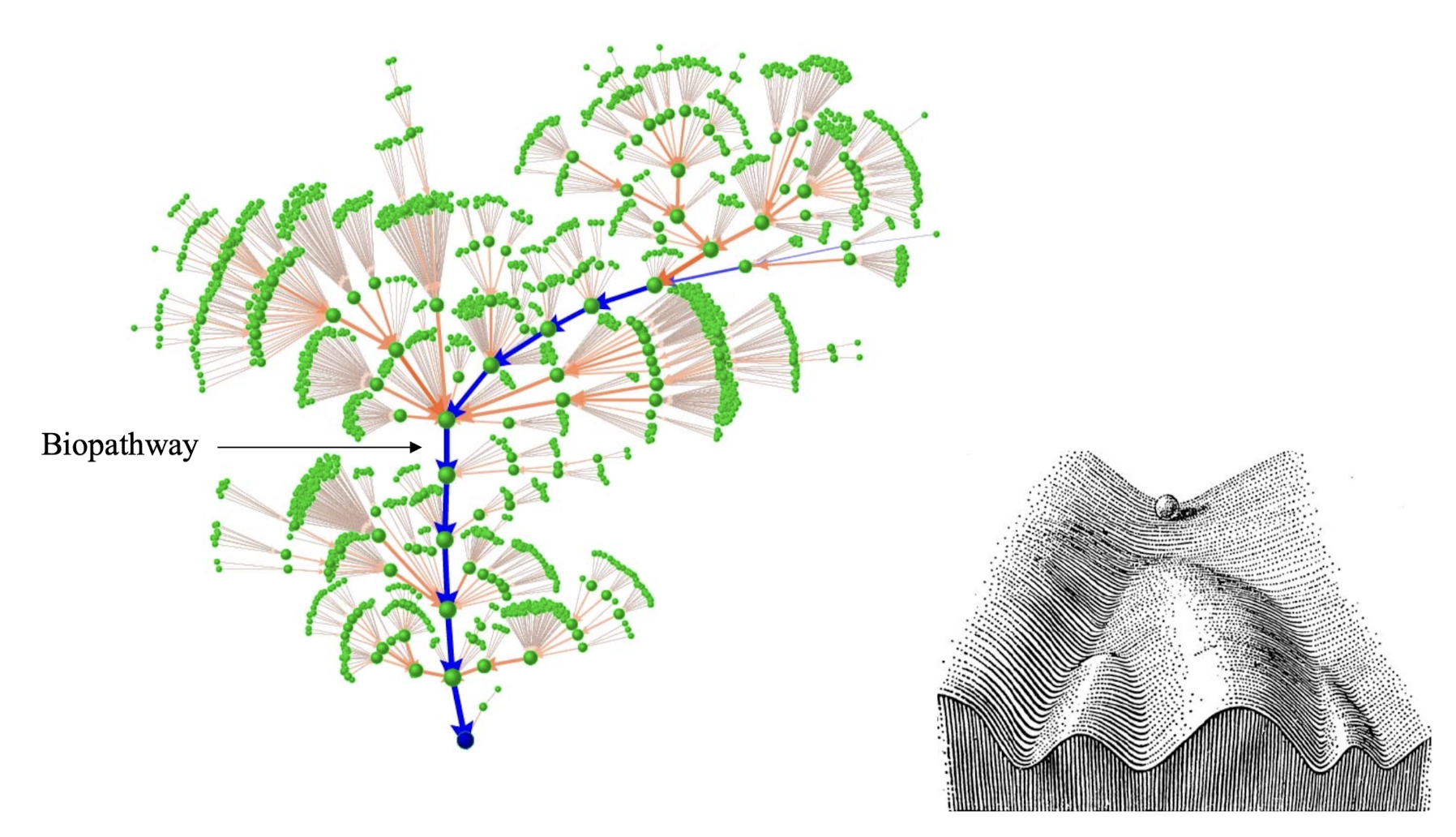
这样的主干道径迹就是一种所谓的吸引子,对于生物而言,甚至可以是一个吸引盆,生物网络的稳定性是非常好的. 这种稳健的特性本质上来源于网络自身的连接方式.
遗憾的是这样的理论取得的成果并不好... 因为我们并没有通过这个理论来解答很多实际问题,这是一个悲伤的故事;不过这样的事情也是常见的.
Hopfield Network & Spin-glass (自旋玻璃):
注意
物理学家在做什么,什么就是物理.
Hopfield 一开始仅仅是组建了一个二十多人的 seminar 团队而已,研究有没有什么 toy model 能够解释「记忆是如何储存在神经网络中的」. 这里面有神经科学家、动物行为学家,还有凝聚态物理学家... 非常混乱的一个团队. 但是也在这种情况下诞生了 Nobel 级别的理论.
自旋玻璃在某种意义上和 Ising 模型是类似的.
本学期课程的最后一个板块 —— 自组织.
更新日志
2025/12/23 13:26
查看所有更新日志
0fcc8-于